Charles, patient homme de 15ans se présente à l'hôpital pour toux persistante, douleurs thoraciques et dyspnée progressive.
L'examen radiologique révèle la présence d'opacités cotonneuses étendues, bilatérales.
Une bronchoscopie + biopsie broncho-pulmonaire révèle la présence d'Aspergillus fumigatus, confirmé à la culture.
Au laboratoire, la formule leucocytaire est normale, avec une répartition normale des leucocytes. Les taux d'imunoglobulines sont normaux. La fonction des phagocytes circulants est pathologique au test de réduction du NBT.
On hospitalise le patient et on le traite par un antimycotique, l'amphotéricine B. Pendant ce temps le patient contracte une infection respiratoire à Pseudomonas aeruginosa ainsi qu'à Streptococcus faecalis. On le traite avec des antibiotiques. L'état du patient s'améliore et il quitte l'hôpital après deux mois.
famille du patient
Le patient a deux frères et deux soeurs. Un de ses frères, Albert, a développé un abcès périnéal à 9ans, et possède aussi des leucocytes ne réduisant pas le NBT. Sa mère et une de ses soeurs ont des PMN mixtes dont certains ne réduisent pas le NBT , d'autres oui.
Des examens approfondis montrent que le patient et son frère de 9ans ont une réduction de la production par les granulocytes de H2O2 (peroxyde d'hydrogène). On leur donne un traitement par IFN-gamma
Composition du sang[]
composition du sang
Le sang (volume total de 5-6L chez l'adulte) est un tissuconjonctif composé de plasma et de cellules. Si on centrifuge du sang en présence d'anti-coagulants, on peut séparer les différents composants. Tout au fond, les érythrocytes remplissent à eux seuls 48% du volume sanguin (38% chez la femme), c'est aussi ce qu'on appelle l'hématocrite. En-dessus se trouve le buffy coat, composé des leucocytes et des plaquettes. Le reste, c'est le plasma, qui contient des sels et des composants organiques (lipides, protéines, acides aminés, vitamines, hormones,...)
Si on n'ajoute pas d'anticoagulant, le sang va se séparer en un caillot (composé des cellules sanguines liées à de la fibrine) et du sérum (contenant le reste, avec absence de fibrinogène)
Globules rouges[]
Les globules rouges, ou érythrocytes, sont des cellules anucléées, biconcaves, constituées d'hémoglobine et d'enzymes glycolytiques. Ils permettent le transport de l'oxygène et du CO2 et leur durée de vie est d'environ 120j. Ils sont éliminés par phagocytose ou détruits par hémolyse dans la rate. Ils sont remplacés par des réticulocytes qui maturent dans les deux premiers jours suivant leur entrée dans la circulation (c'est 1-2% des globules rouges circulants).
Leucocytes: granulocytes, agranulocytes[]
Les leucocytes sont soit des granulocytes (présence de granules primaires et secondaires), soit des agranulocytes (présence de granules primaires seulement)
Comparés aux érythrocytes et aux plaquettes, les leucocytes semblent peu nombreux, mais on en trouve quand même 6'000 à 10'000 par mm3 dans le sang.
Cellules
Quantité par mm3
Erythrocytes
4 millons - 6 millions
Plaquettes
300'000
Leucocytes
6'000 - 10'000
En réponse à un stimulus, les leucocytes sont capables de quitter la circulation sanguine (par diapédèse) et d'entrer dans les tissus conjonctifs par le mécanisme du homing.
Granulocytes[]
Les granulocytes (ou PMN) possèdent un noyau multilobé et sont capables de phagocyter. Leur cytoplasme contient des granules. Leur durée de vie diffère suivant le type, qu'on peut distinguer suivant leur cytoplasme:
neutrophiles
Les neutrophiles représentent 60-70% des leucocytes. Leur cytoplasme d'aspect rose pâle contient à la fois des granules primaires (contenant des élastases et myélopéroxydases) et secondaires (ou spécifiques, contenant des lyzozymes, NADPH oxydase et protéases). Ils ont une durée de vie de 6-7 heures dans la circulation sanguine, mais peuvent survivre jusqu'à 4 jours dans le tissu conjonctif. Ils quittent la circulation par des veinules post-capillaires et vont éliminerlesbactéries opsonisées ou limiterl'étendue de l'inflammation dans les tissus conjonctifs.
eosinophiles
Les éosinophiles représentent 2-4% des leucocytes. Leur cytoplasme contient de larges granules rouges (contenant de l'éosine peroxydase, de la MBP et de la protéine cationique éosinophile) et leur noyau est bilobé. Comme les neutrophiles, les éosinophiles peuvent quitter la circulation et migrer dans les tissus conjonctifs. Leur rôle est la défense contre les parasites (ce qui n'est pas bactérie, virus et champignon). Ils sont aussi responsables de l'asthme bronchique.
basophiles
Les basophiles représentent 1% des leucocytes. Leur cytoplasme contient de larges granules métachromatiques (contenant des protéines acides sulfatés ou carboxylées, de l'héparine, kallikréine - attire les éosinophiles - et péroxydase) qui obscurcissent souvent le noyau bilobé. Comme les neutrophiles, les basophiles peuvent quitter la circulation et migrer dans les tissus conjonctifs, où ils deviennent similaires à des mastocytes (qui sont toujours dans les tissus), avec capacité de libérer de l'histamine (dégranulation diffuse dans le sang) car ils possèdent un récepteur aux IgE à leur surface. Ils jouent un rôle dans les réactions d'hypersensibilité immédiate (asthme) et retardée (réactions allergiques cutanées), ainsi que dans la propagation de la réponse immunitaire.
Agranulocytes[]
Les agranulocytes sont appelés ainsi car ils ne possèdent que des granules primaires de type lyzosomes (qui sont tellement petits qu'on les voit à peine au microscope, contrairement aux granules secondaires des granulocytes qui sont bien visibles). On en distingue deux types:
lymphocytes
Les lymphocytes représentent 20-40% des leucocytes. Ilspeuvent être petits (97% des cas, = taille d'un GR), ou grands (3% des cas). Ils possèdent un noyau rond (ou un peu encoché), avec un cytoplasme basophile très fin. Les lymphocytes ont une durée de vie allant de quelques jours à quelques années. Les lymphocytes B sont produits et maturent au niveau de la moelle osseuse, et peuvent se différencier en plasmocytes sécrétant des anticorps. Les lymphocytes T sont aussi produits dans la moelle osseuse mais maturent au niveau du thymus. Ce sont les médiateurs de la réponse immunitaire à médiation cellulaire.
monocytes
Les monocytes représentent 2-8% des leucocytes. Ils sont plus grands que les lymphocytes et possèdent un noyau en forme de rein (ou ovale). Ils circulent dans le sang pendant 12-100h puis passent dans le tissu conjonctif où ils se différencient en macrophages (avec fonction de phagocytose, présentation d'antigène et nettoyage des débris de cellules mortes). Dans les os, les monocytes se différencient en ostéoclastes, sous contrôle des ostéoblastes.
Leucocytes
Proportion
Neutrophiles
60-70%
Lymphocytes
20-40%
Monocytes
2-8%
Éosinophiles
2-4%
Basophiles
1%
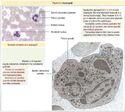
Plaquettes[]
plaquettes
Les plaquettes sont des petits fragments de cytoplasme dérivés des mégacaryocytes, sous contrôle de la TPO produite par le foie et les reins. Les mégacaryocytes développent des projections cytoplasmiques qui deviennent des proplaquettes, dont la fragmentation forme les plaquettes. Ce processus de différentiation prend 10-12 jours. Les Plaquettes sont capables de se lier à la TPO et de la dégrader, ce qui permet une régulation de leur propre production.
La membrane plasmique des plaquettes s'invagine à plusieurs endroits pour former un système canaliculaire ouvert. La région centrale des plaquettes s'appelle le granulomère (qui contient notamment les granulations), tandis que la périphérie s'appelle le hyalomère et contient des microtubules et microfilaments pouvant modifier la forme de la plaquette.
Hématopoïèse et cellules hématopoïétiques[]
L'hématopoïèse commence chez le fœtus pendant le premier trimestre, par des îlots hématopoïétique du sac vitellin. Les îlots proviennent des hémangioblastes (progéniteurs des cellules hématopoiétiques et endothéliales). L'hématopoïèse continue pendant le deuxième trimestre dans le foie puis dans la rate. Au 7e mois, la moelle osseuse finit par devenir le site primaire d'hématopoïèse, et restera ainsi pendant la vie adulte. On trouve chez un adulte 1 milliard de cellules hématopoïétiques pour 1,7ml de moelle osseuse.
moelle osseuse
On trouve des cellules endothéliales, de macrophages et de vaisseaux sanguins, entremêlés à l'intérieur des travées osseuses. Les fibroblastes, les cellules de soutien et les cellules endothéliales produisent des facteurs de croissance hématopoïétiques et des cytokines régulant la production de cellules sanguines.
cellule
fonction
adipocytes
source locale d'énergie, synthèse de facteurs de croissance
fibroblastes
régulation de l'hématopoïèse
cellules conjonctives de soutien
régulation de l'hématopoïèse
cellules vasculaires endothéliales
régulation de l'hématopoïèse, barrière empêchant les cellules immatures de quitter la moelle et permettant aux cellules matures d'entrer dans le sang
macrophages
élimination des cellules apoptotiques, des débris, protection contre les substances étrangères
structure moelle
Le compartiment cellulaire hématopoïétique fournit la quantité et les types cellulaires adaptés aux besoins physiologiques. Les cellules hématopoïétiques sont regroupées dans des "niches" dans la moelle osseuse et possèdent des capacités d'auto-renouvellement, de différenciation, de croissance, et de maturations en différents types cellulaires.
Le compartiment cellulaire hématopoïétique est très vascularisé, via l'artère longitudinale centrale, provenant elle-même de l'artère nourricière. L'artère longitudinale centrale donne les plexus capillaires médullaires. Ces plexus sont drainés par les sinus veineux médullaires, qui se jettent dans la veine longitudinale centrale, qui se jette finalement dans la veine nourricière.
Les cellules hématopoïétiques matures migrent depuis leur lieu de croissance dans les sinusoïdes par un processus de migration transendothéliale actif à travers les fenestrations des sinusoïdes, avant de rejoindre la circulation via la veine nourricière.
Les cellules hématopoïétiques immatures n'ont pas la capacité de migration transendothéliale, et sont retenues dans leur compartiment par les cellules endothéliales (qui sont des cellules spécialisées capable de phagocytose ainsi que de production de facteurs de croissance stimulant la prolifération et différenciation des cellules hématopoïétiques).
hématopoïèse référence
Les cellules souches hématopoïétiques CD34+ donnent naissance à toutes les lignées. Grâce aux facteurs FLT3-L et SCF, elles peuvent donner les différents précurseurs.
Précurseur
Va donner
Via
Myéloïde
Érythrocyte
EPO, (IL-3, SCF)
Megacaryocyte
TPO, (IL-3)
Monocyte (CFU-M)
Macrophage
Dendrocyte
Ostéoclaste
IL3, GM-CSF, M-CSF,
M-CSF
(GM-CSF, IL-4)
RANK-L
PMN (CFU-G)
Neutrophile
Éosinophile
Basophile
IL3, G-CSF
G-CSF
IL-5
IL-4
Lymphoïde
Lymphocyte B
IL-11+IL-7, IL-6
Lymphocyte T
IL-1+IL-7, IL-2
NK
NK
IL-2,IL-15
Mastocyte
Mastocyte
SCF
La moelle osseuse est composée de trois populations principales: les cellulessoucheshématopoïétiques (capables d'auto-renouvellement - cellules multipotentes produisant les cellules souches myéloïdes et lymphoïdes), les cellulesprogénitricesengagées (donnant les différentes lignées cellulaires - cellules souches myéloïdes et lymphoïdes) et les cellules en voie de maturation ou précurseur (à l'origine des cellules circulantes).
Érythropoïèse[]
L'érythropoïèse comprend la séquence suivante: proérythroblaste, érythroblaste basophile, érythroblaste polychromatophile, érythroblaste otochromatique (ou acidophile, qui expulse son noyau qui est ensuite mangé par un macrophage), réticulocyte (qui se transforment en érythrocyte en 24h une fois dans la circulation), érythrocyte
Erythropoïèse
Le régulateur majeur de l'érythropoïèse est l'EPO, une glycoprotéine produite (à 90%) au niveau des reins, par les cellules juxtaglomérulaires interstitielles du cortex. La production d'EPO est stimulée par l'hypoxie, que les cellules juxtaglomérulaires interstitielles détectent via la prolyl hydroylase dépendante de l’oxygène (une protéine qui inhibe la production du facteur de transcription HIF-1alpha). Lorsque les niveaux d'oxygène sont bas, la prolyl hydroxylase dépendante de l'oxygène devient inactive, et le HIF-1alpha non hydroxylé peut produire de l'EPO.
L'EPO va agir en stimulant la prolifération des cellules érythroïdes progénitrices.
En cas d'insuffisance rénale, on peut donner de l'EPO-recombinante. On peut voir l'efficacité du traitement en mesurant une augmentation des réticulocytes circulants.
L'érythroblastepolychromatophile est indépendant de l'EPO et mitotiquement actif, c'est lui qui est responsable spécifiquement de la synthèsedel'hémoglobine. Les cellules qui suivent (érythroblaste otochromatique, réticulocytes et érythrocytes) n'ont pas d'activité mitotique.
Leucopoïèse: granulocytes, agranulocytes[]
La leucopoïèse comprend la formation des granulocytes et des agranulocytes.
Granulocytse[]
lignée myéloïde
La lignée des granulocytes fait partie de la lignée des cellules souche myéloïdes et comprend la séquence suivante: myéloblaste, promyélocyte, myélocyte, métamyélocyte, band cell (polynucléaire à noyau peu segmenté), forme mature
Par cette lignée seront créés les neutrophiles (via G-CSF ; prennent 10-14j pour apparaître en condition normale et moins dans une situation infectieuse), les éosinophiles (via IL-5) et les basophiles (via IL-4).
Les myéloblastes, promyélocytes et myélocytes sont mitotiquement actifs. Les métamyélocytes et les band cells n'ont pas d'activité mitotique, mais continuent à se différencier. Un processus typique de la maturation est l'apparition des granules primaires et secondaires. Les myéloblastes n'ont pas de granules. Les granules primaires apparaissent chez les promyélocytes, et les granules secondaires chez les myélocytes.
Agranulocytes[]
Les agranulocytes comprennent les lymphocytes et les monocytes.
Les lymphocytessont très diversifiés quant à leur origine, durée de vie, localisation dans les organes lymphatiques, marqueurs à la surface et fonction.
La cellule souche lymphoïde peut donner des progéniteurs des cellules B et des cellules T. Les lymphocytes B maturent dans la moelle osseuse, puis migrent dans les organes lymphoïdes. Les cellules T maturent dans le thymus, puis migrent dans les organes lymphoïdes.
Les précurseurs des lymphocytes s'appellent les lymphoblastes, qui deviennent prolymphocytes puis lymphocytes.
monocytes macrophages
Les monocytes proviennent des CFU-M. La lignée des monocytes comprend la séquence suivante: monoblaste (mitotiquement actif), promonocyte (mitotiquement actif), monocyte (contient des granulations de lysosomes primaires et des petites vacuoles), macrophage
Quand le monocyte quitte la circulation, il devient macrophage. On en trouve dans une grande variété de tissus (poumons, tractus digestif, rate, foie, péritoine, nœuds lymphatiques, os,...). Au niveau de la peau, les monocytes donnent les cellules de Langerhans. Au niveau niveau du foie, ils donnent les cellules de Kupffer.
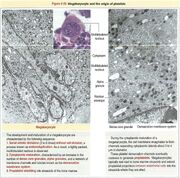
Plaquettes et mégacaryocytes[]
megacaryocytes plaquettes
Les plaquettes(ou thrombocytes) proviennent du mégacaryoblaste, lui-même dérivé de la cellule souche myéloïde. Le mégacaryoblaste va donner le promégacaryocyte, puis le mégacaryocyte. Histologiquement, on le confond souvent avec les ostéoclastes car les deux sont des grosses cellules.
Le mégacaryocyte possède un noyau irrégulièrement segmenté à la suite de plusieurs processus de division endomitotique (mitose sans division cellulaire). Sa membrane forme des invaginations qui vont donner les proplaquettes, qui se fragmenteront en plaquettes.
Barrières contre les infections[]
Barrières physiques
La peau est l'organe servant de première défense pour empêcher l'entrée des pathogènes, grâce à ses nombreuses couches d'épithélium ainsi que la kératine à la surface. La barrière de la peau peut être rompue par différents types de dégâts (blessures, brûlures, chirurgie,...) qui la rendent vulnérable.
Les muqueuses respiratoire, pulmonaire et gastrointestinale sont en continuité avec la peau et sont plus fragiles. Elles sont tapissées de mucus qui protège l'épithélium et limite les infections. Au niveau des poumons, le mucus est constamment renouvelé et évacué par des cellules ciliées.
Les surfaces épithéliales sécrètent aussi des substances antibactériennes. Les glandes sébacées sécrètent du sébum contenant des acides gras et de l'acide lactique qui inhibent la croissance bactérienne à la surface de la cellule. Les épithéliums produisent aussi de la défensine qui tue les bactéries, champignons et virus enveloppés en perturbant leur membrane. Les microorganismes sont aussi tués par l'environnement acide de l'estomac, du vagin et de la peau.
Avec ces barrières, la majorité des pathogènes est repoussée (c'est la première défense contre les pathogènes).
Réponse immunitaire innée[]
En général, lorsqu'un pathogène passe les barrières physiques, les infections sont très localisées et sont résolues en quelques jours sans provoquer la moindre maladie. De telles infections sont contrôlées rapidement grâce au système immunitaire inné, qui est toujours prêt à agir. Son action se fait en deux temps:
reconnaissance pathogène
D'abord, le pathogène doit être reconnu. Cela ce fait par liaison de protéines solubles ou de récepteurs membranaires soit sur le pathogène et ses produits, soit sur cellules infectées par le pathogène ou des protéines sériques altérées par le pathogène.
Ensuite, lorsque le pathogène est reconnu, des mécanismes vont être mis en place pour le détruire, via des cellules effectrices. Les bactéries se font phagocyter, les cellules infectées se font tuer, les parasites attaquer, etc. Pour renforcer l'action, le système du complément aide les cellules effectrices en opsonisant les pathogènes et en les attaquant de lui-même (le complément est composé de deux parties, une qui reste sur liée de façon covalente sur le pathogène et le marque comme dangereux et une autre soluble qui va recruter des cellules effectrices).
Cette immunité est appelée innée car on l'hérite de nos gènes.
Les récepteursprotéiques reconnaissant les pathogènes sont assez variés, et se lient à divers ligands (peptides, protéines, glycoprotéines, protéoglycans, petidoglycans, hydrates de carbone, glycolipides, phospholipides et acides nucléiques).
inflammation
La réponse générale du système immunitaire inné est de produire une inflammation au niveau du tissus infecté. L'inflammation se caractérise par la rubor, dolor, calor, tumor. Ce n'est donc pas l'infection elle-même mais la réponse à l'infection qui fait l'inflammation.
migration leucocytes
Ces effets vont être déclenchés par les cytokines que relâchent les cellules effectrices lorsqu'elles reconnaissent le pathogène pour déclencher la réponse immunitaire innée. Les cytokines vont induire une vasodilatation locale (rougeur et chaleur), avec augmentation de la perméabilité vasculaire (œdème). L’œdème va faire pression sur les terminaisons nerveuses libres (douleur).
Les cytokines changent aussi les propriétés de l'endothélium, permettant à des leucocytes de s'y attacher et de faire une migration trans-endothéliale (rolling par interaction des sélectines de l'endothélium sur les ligands aux sélectines du leucocyte (Sialyl LewisX) puis binding par adhésion des intégrines LFA-1 du leucocyte sur les ICAM de l'endothélium - ce processus est stimulé par la cytokine CXCL8 (= IL-8), qui augmente l'interaction et les cytokines qui permettent l'expression des molécules de surface de l'endothélium et du leucocyte). Une fois arrivées dans les tissus enflammés, ces cellules inflammatoires vont sécréter des substances qui vont contribuer à l'inflammation, à la douleur et à l’œdème. Le but de l'inflammation est donc de permettre l'arrivée rapide et en grand nombre des cellules immunitaires.
Réponse immunitaire adaptative[]
innee vs adaptative
En générale, l'immunité innée s'occupe de la majorité des infections. Mais certaines infections outrepassent ce système (surtout chez les personnes fatiguées, stressées, mal nourries,...). Dans ces cas, la réponse innée ne peut que ralentir la progression de l'infection, tout en appelant en renfort la réponse adaptative, c'est-à-dire les lymphocytes (qui augmentent la spécificité et la puissance de la réponse). La réponse adaptative produit une immunité longue-durée contre un pathogène particulier.
selection lymphocyte
Contrairement aux récepteurs de l'immunité innée qui sont d'une grande variété, mais pas contre un pathogène particulier, les récepteurs de l'immunité adaptative sont du même type cellulaire, mais hautement spécifique à un seul pathogène. Ils sont codés par des gènes qui sont coupés, réarrangés et épissés, afin de créer aléatoirement des milliards de combinaisons aléatoires. Chaque lymphocyte possède une variante différente.
Lors de l'infection, seul les lymphocytes avec une variante efficace vont être sélectionnés et participer à la réponse adaptative. Ces lymphocytes vont proliférer et se différencier pour produire une grande quantité de cellules effectrices spécifiques contre le pathogène en question. Ces processus sont appelés sélectionclonale et expansionclonale, et prennent environ une semaine à se mettre en place (la grippe par exemple, dure environ une semaine, le temps que la réponse adaptative se mette en place).
Certains lymphocytes sélectionnés durant l'infection persistent dans le corps et permettent une mémoire immunologique à long terme (ou immunitéacquise ou protectrice), via la formation de cellules mémoire à longue vie. Si une nouvelle infection du pathogène survient, la réponse sera plus rapide et plus forte que la première fois et peut ne pas laisser le temps au pathogène d'induire la maladie (réponse immunitaire secondaire - contrairement à la réponse immunitaire primaire, qui est la première stimulation du système immunitaire adaptatif). Pour certains pathogène, une infection suffit pour donner une mémoire immunologique durant plus d'une dizaine d'années. Le principe des vaccins est d'induire une telle immunité (l'immunité due à la vaccination nécessite une réponse innée et acquise, car la réponse adaptative est déclenchée par la réponse innée).
deficit inne ou adaptatif
L'immunité adaptative est mieux comprise que l'immunité innée. La médecine s'occupe principalement des infections qui outrepassent les capacités de l'immunité innée, comme les pneumonies, la rougeole, ou la grippe, et qui stimulent l'immunité adaptative.
Cellules du système immunitaire[]
cellules hematopoietiques
Toutes les cellules du système immunitaire sont dérivées des cellules souches hématopoïétiques, via l'hématopoïèse (qui se fait en continu). Chez l'adulte, l'hématopoïèse se fait principalement dans la moelle osseuse des os du crâne, des côtes, du sternum, de la colonne vertébrale, du bassin et du fémur. Les cellules hématopoïétiques possèdent une durée de vie assez courte et doivent être constamment renouvelées.
Les érythrocytes sont les globules rouges transportant l'oxygène.
Les mégacaryocytes (qui résident dans la moelle osseuse) donnent les plaquettes, qui sont des petits fragments cellulaires sans noyau. Les plaquettes participe à la formation du caillot lorsqu'un vaisseau sanguin est endommagé.
action neutrophiles
Les neutrophiles sont spécialisés dans la phagocytose des pathogènes. Ce sont les plus nombreux et les plus létaux des phagocytes. Dans la moelle osseuse, 60% de l'hématopoïèse est dédiée à leur production. En cas d'infection, ils sont rapidement mobilisés sur le site et sont efficaces même en milieu anaérobique (condition souvent présente sur un site d'infection). Ils sont stockés dans la moelle osseuse et ont une courte vie (24-48h). Ils meurent sur le site d'infection, où ils sont phagocytés par le macrophages. Le pus est formé de neutrophiles morts.
Les éosinophiles sont bien moins nombreux. Ils servent à la défense contre les vers et autres parasitesintestinaux.
Les basophiles sont le moins nombreux. Ils sont aussi utiles contre des parasites, mais entrent très rarement en action.
macrophages vs pathogene
Les monocytes circulent dans le sang et migrent dans les tissus, pour se différencier en macrophages. Les macrophages peuvent phagocyter, comme les neutrophiles. Ils sont un peu les éboueurs du corps, phagocytant et se débarrassant des cellules mortes et des microorganismes invasifs. Contrairement aux neutrophiles, les macrophages ont une longue durée de vie. Ce sont souvent les premières cellules à entrer en contact avec le pathogène, et les signaux qu'ils envoient aux autres cellules leur donnent un rôle de chef d'orchestre (ils attirent notamment les neutrophiles et autres leucocytes et activent les cellules T).
Les cellules dendritiques résident dans les tissus. Elles ressemblent fortement aux macrophages, mais possèdent la fonction unique d'agir comme messagers cellulaires envoyés pour demander de l'aide à la réponse adaptative quand cela est nécessaire (notamment via l'activation des cellules T). En effet, les cellules dendritiques sont capables de quitter le tissus infecté et de migrer jusqu'aux organes lymphoïdes, en emportant avec elles des cargaisons de pathogènes intacts et dégradés.
Les mastocytes résident dans les tissus conjonctifs et ressemblent au basophiles (en plus grand), mais leur dégranulation (histamine, sérotonines) est extracellulaire. Ils possèdent un récepteur aux IgE à leur surface. Leur dégranulation sur les sites d'infections contribuent pour beaucoup à l'inflammation.
Les cellules NK (Natural Killers, ou "grands" lymphocytes) font partie de la réponse immunitaire innée. Ils sont importants dans la défense contre les infections virales. Ils entrent dans les tissus infectés et limitent l'infection virale en tuant les cellules infectées et en sécrétant des cytotoxines qui entravent la réplication virale dans les cellules (perforin, granzyme).
Différenciation cellule B en plasmocyte
Les lymphocytes (ou "petits" lymphocytes), sont responsables de la réponse adaptative. Ils sont petits car ils circulent sous forme immature quiescente dans la circulation. La reconnaissance d'un pathogène par un lymphocyte lance un processus de différenciation et de maturation, permettant une réponse puissante dirigée contre le pathogène, le tout en l'espace de 1-2 semaines. Les lymphocytes peuvent être divisés en lymphocytes B (dont le récepteur de surface est une immunoglobuline spécifique) et lymphocytes T (qui possède un T-cell receptor spécifique). Les lymphocytes T peuvent être subdivisés en cytotoxiques et en helper. Les lymphocytes T cytotoxiques tuent les cellules infectées par des virus ou des bactéries intracellulaires, tandis que les helper sécrètent des cytokines qui permettent aux autres cellules immunitaires de s'activer pleinement (par exemple, ils activent les lymphocytes B en plasmocytes). Les plasmocytes sécrètent les AC (des IG solubles) dans la circulation.
Les cellules souches hématopoïétiques peuvent se diviser pour donner de nouvelles cellules souches (self renewal) ou peuvent se différencier en cellules souches plus matures, qui peuvent donner trois lignées différentes:
- érythroïde (érythrocytes et mégacaryocytes)
- myéloïde (granulocytes ou PMN - neutrophiles, basophiles et éosinophiles - ainsi que mastocytes et monocytes - macrophages et cellules dendritiques)
- lymphoïde (cellules NK et lymphocytes)
Tissus lymphoïdes[]
tissus lymphoides
La majorité des lymphocytes se situent au niveau des tissus lymphoïdes (ou organes lymphoïdes). Ils sont répartis dans plusieurs lieux différents:
La moelle osseuse
Le thymus
La rate
Les amygdales
L'appendice
Les ganglions lymphatiques
Les plaques de Peyer
Les muqueuses respiratoire, digestive, urogénitale (tissu lymphoïde moins organisé)
Les tissus lymphoïdes peuvent être classés en deux types: primaire et secondaire.
Les tissus lymphoïdes primaires(ou centraux) sont les lieux de développement et de maturation des lymphocytes. Il s'agit de la moelleosseuse (lymphocytes B et T notamment - les lymphocytes B maturent dans la moelle) et du thymus (lieu de maturation des lymphocytes T).
recirculation lymphocytes
Les tissus lymphoïdes secondaires (ou périphériques) comprennent tous les autres tissus lymphoïdes. Ils correspondent aux lieux ou les lymphocytes matures sont stimulés et répondent à l'invasion des pathogènes. Les nœuds lymphatiques se trouve à la jonction des vaisseaux lymphatiques (qui prennent leur origine dans les tissus conjonctifs et drainent le plasma). La lymphe retourne dans la circulation sanguine via le canal thoracique qui se jette dans la veine sous-clavière gauche. Le flux de la lymphe est assez lent car il n'y a pas de pompe dédiée à la faire circuler. Son flux est unidirectionnel, grâce à la présence de valves. C'est le fait de bouger le corps qui permet de la faire circuler, ainsi si un patient est immobile, son flux lymphatique diminue, provoquant des œdèmes. La lymphe est aussi le lieu de déplacement des lymphocytes B et T (ce sont les seules cellules qui s'y trouvent). Les lymphocytes peuvent rejoindre la lymphe via les capillaires irriguant les tissus lymphoïdes. Sauf activation, les lymphocytes passent quelques temps dans les nœuds lymphatiques puis circulent de nouveau (à un moment donné, la majorité des lymphocytes se trouve cependant dans les organes lymphatiques et les tissus lymphoïde). Ces mouvements de circulation des lymphocytes sont appelés la "recirculation des lymphocytes". La rate par contre n'a aucune connexion avec la lymphe (ainsi, les lymphocytes entrent et sortent de la rate via la circulation sanguine).
lieu de rencontre
Le lieu de rencontre entre le lymphocyte et le pathogène sera un tissu lymphoïde secondaire. En principe, les pathogènes vont infecter un tissu conjonctif (ils y entrent lors d'une lésion). De cet endroit, les pathogènes, des parties des pathogènes ou des cellules dendritiques transportant des pathogènes vont être transportés par les vaisseaux lymphatiques afférents jusqu'à un ganglion lymphatique à proximité (draining lymph node), où les cellules dendritiques s'arrêtent pour présenter le matériel aux lymphocytes passant par là (et les pathogènes détruits par les macrophages). Si le lymphocyte reconnait le pathogène et s'active, il va proliférer, ce qui fera gonfler le ganglion.
La rate[]
Les pathogènes peuvent entrer directement via le sang (si on se fait piquer par un moustique par exemple). La rate est un organe permettant de filtrer le sang. Un de ses rôles est de nettoyer le sang des globules rouges endommagés ou sénescents (dans la pulpe rouge). L'autre rôle essentiel est de lutter contre les infections apparaissant directement dans le sang (au niveau de la pulpe blanche). En effet une infection sanguine est un risque d'infection systémique. Les pathogènes sont captés dans la rate par des macrophages et des cellules dendritiques, puis sont présentés à des lymphocytes arrivant par le sang (au niveau de la pulpe blanche).
Certains individus naissent sans rate (asplénie), et sont susceptibles à certaines infections à bactéries encapsulées. Lors de dommage accidentel à la rate, il est parfois indispensable de la retirer. Cette condition n'est en général pas très grave, mais on fait en général quelques vaccins préventifs.
Tissus lymphoïde digestif[]
Le tractus digestif est l'endroit du corps où l'on trouve la plus grande variété et la plus grande quantité de pathogènes, avec le système respiratoire. Leurs muqueuses sont donc très à risque et c'est pour ça qu'elles sont renforcées au niveau digestif par des GALT, des amygdales, des plaques de Peyer et l'appendice, et au niveau respiratoire par des BALT. En général, les tissus lymphoïdes diffus dans les muqueuses s’appellent des MALT.
Les pathogènes rejoignent les MALT directement depuis la muqueuses grâce à un transport par les cellules M.
Différents pathogènes[]
Les pathogènes peuvent provoquer des infectionsintracellulaires ou des infectionsextracellulaires. Cela a comme conséquence que les mécanismes pour lutter contre ces pathogènes seront différents suivant leur localisation (les mécanismes de défense solubles ne peuvent pas atteindre les infections intracellulaires, mais agissent sur les infections extracellulaires, mais on peut tuer les cellules qui sont infectées).
La majorité des pathogènes ne sont capable d'infecter qu'un petit nombre d'hôtes différents, ainsi il est rare pour un homme de contracter une infection de la part d'un autre animal. La majorité des infections se feront donc par transmission d'homme à homme, soit directement, soit indirectement via un passage par une autre espèce pour compléter le cycle du pathogène.
Complément[]
Une des premières armes qui attaque les pathogènes lors d'infections est le système du complément. Il s'agit d'un réseau de protéines solubles produites par le foie, qui circulent dans le sang et la lymphe, ainsi que dans le liquide extra-cellulaire. Le complément agit en série, afin de faciliter l'élimination des pathogènes.
roles du complement
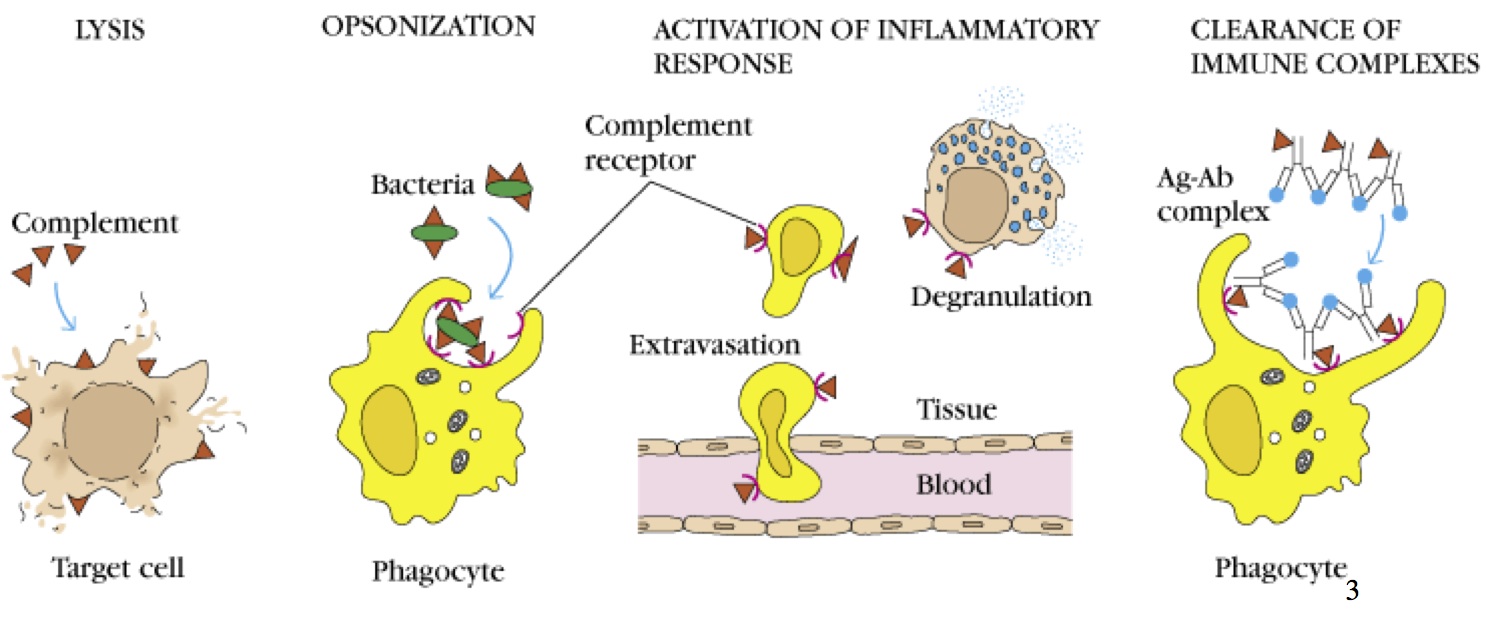
Les rôles du complément sont
Lyse directe du pathogène
Opsonisation (facilitation de la phagocytose en formant un manteau sur le pathogène)
Activation de la réponse inflammatoire
Facilitation de l'extravasation
Dégranulation des mastocytes (C5aR)
Nettoyage des complexes immuns et des cellules apoptotiques
(Augmentation de la production d'anticorps)
(Participation dans l'établissement des cellules mémoires)
Le complément est composé d'enzymes (ou protéases) qui circulent sous forme inactivée (ou zymogènes). L'infection provoquera l'activation du complément, avec une série de cascades de protéases, dont la plus importante est la C3 (une déficience en C3 rend vulnérable à de sévères infections). En effet, toute infection aboutit au clivage de C3 (produit par le foie) en C3a et C3b (ce dernier se lie de façon covalente sur le pathogène - on parle de fixation du complément). La liaison de C3b sur le pathogène le marque comme dangereux et permet d'endommager la membrane. Le C3a permet de recruter les cellules du système immunitaire par chémoattraction.
voies du complement
Le complément comporte trois voies: la voie alternative, la voie des lectines et la voie classique.
La voie qui agit au début d'une infection est la voie alternative, c'est aussi la voie la plus importante, surtout pour les infections bactériennes.
La voiedeslectines fait aussi partie de l'immunité innée mais nécessite du temps avant de fonctionner.
La voieclassique est à l'interface entre immunité innée et adaptative car elle peut démarrer soit via la fixation d'AC (immunité adaptative), soit via la fixation de la CRP (immunité innée).
Au niveau de l'évolution, la voie alternative fut la première à apparaître, suivie par la voie des lectines puis finalement par la voie classique.
Voie classique[]
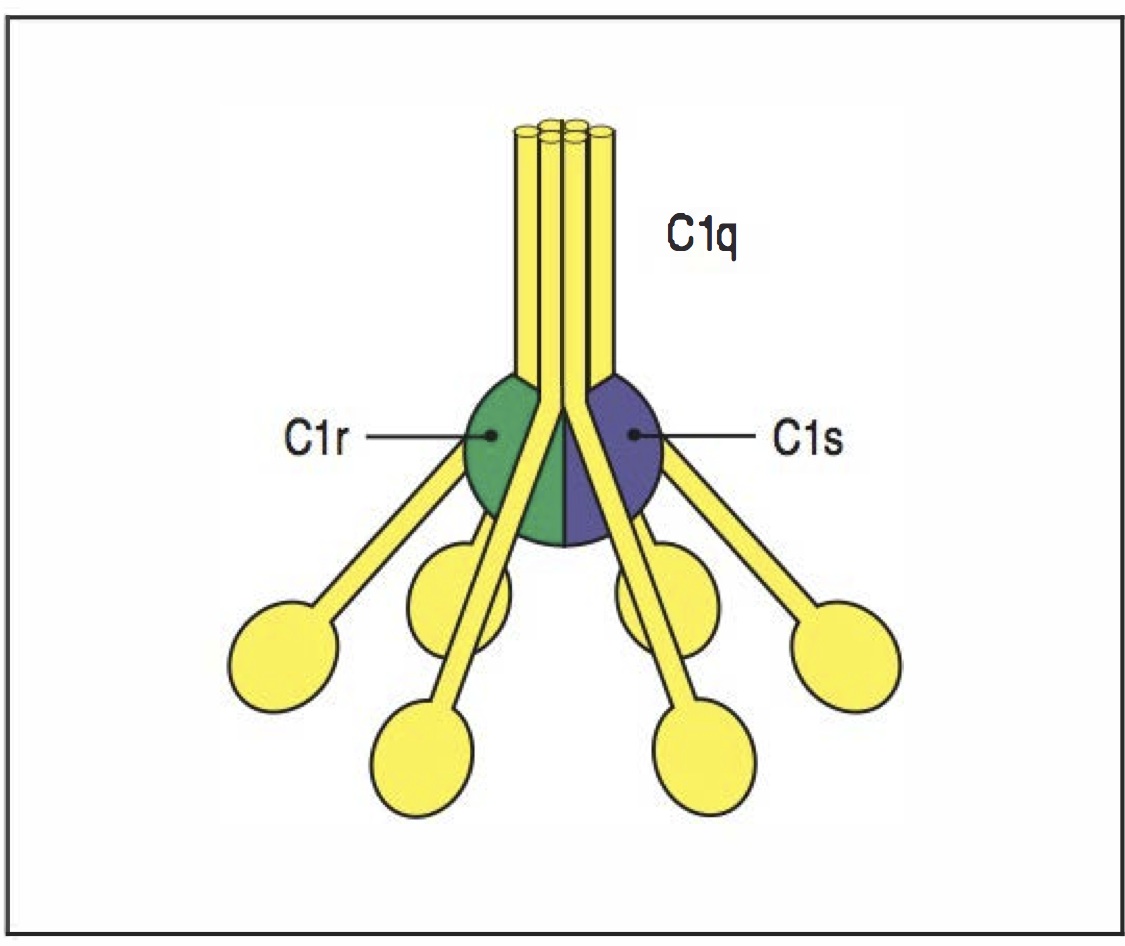
structure C1
La voie classique commence par la collectine C1, composée de C1q ainsi que de C1r et C1s (qui ne sont actives que quand C1q est liée). La C1q peut se lier soit à une Ig fixé sur un antigène à la surface du pathogène (immunité adaptative), soit à la CRP, soit sur des ligandsdecellulesapoptotiques .
activation C1q
Les anticorps IgM & IgG peuvent activer la voie classique du complément.
1) IgM
Les IgM se trouvent toujours dans le sang sous forme pentamérique formant ainsi une structure plane composée par 5 IgM reliées entre elles par des ponts dissulfures ressemblant à un flocon de neige.
Les parties hypervariables des 5 IgM reconnaissent le peptide antigénique présenté à la surface d'une cellule infectée par le pathogène pour lequel elles sont spécifiques ce qui leur permet d'adopter une conformation dite en « agrafe ». Cette conformation en agrafe se lie très bien à la partie C1q du complément ce qui entraîne l'activation de la voie classique du complément.
2) IgG
Les IgG sont toujours sous forme monomérique et peuvent se trouver dans le sang ou dans le fluide extracellulaire. Les anticorps IgG peuvent également activer la voie classique du complément. Pour ce faire, il faut qu'il y ait au minimum 2 anticorps IgG qui reconnaissent l'antigène présent à la surface d'une cellule ou sur l'antigène soluble. Une fois que 2 IgG se sont liés au peptide antigénique pour lequel ils sont spécifiques, ces derniers se lient à la molécule C1q du complément ce qui entraîne l'activation de la voie classique du complément.
Il faut donc que C1q soit lié à plusieurs IgG à la surface du pathogène pour pouvoir correctement s'activer.
Ig
Structure
Sites de liaison à C1q
Activateur de C1q
IgM
pentamère
3
efficace
IgG
monomère
1
moins efficace
voie classique
La cascade de réactions qui s'ensuit est la suivante:
C1r et C1s vont cliver le C4 en C4a et C4b, ainsi que le C2 en C2a et C2b.
C4b s'associe avec C2a pour former C4b2a (une C3 convertase)
C4b2a clive jusqu'à 1'000 C3 en C3a et C3b.
C3b s'associe avec C4b2a pour former C4b2a3b (une C5 convertase). Il peut aussi se lier tout seul à la surface du pathogène
thioester C3b
Le clivage de C3 en C3a et C3b par la C3 convertase expose à l'environnement une liaison thioester sur le C3b. Cette liaison peut se faire spontanément hydrolyser par l'eau, mais certaines C3b au lieu de ça vont réagir avec la surface du pathogène et s'y attacher.
C'est grâce à ce mécanisme que C3b peut se lier de manière covalente avec les pathogènes.
Une fois formée, la C5 convertase va pouvoir cliver C5 en C5a et C5b, étape commune aux trois voies.
structure MBL
voie des lectines
Voie des lectines[]
La voie des lectines débute par la liaison de la MBL(une collectine, et aussi un PRR) sur un résidu mannose à la surface du pathogène. La MBL ressemble au C1q et possède aussi deux enzymes avec la même fonction que C1r et C1s, il s'agit de MASP-1 et MASP-2.
La voie des lectines ressemblent beaucoup à la voie classique, et implique la suite de réactions suivantes:
MASP-2 va cliver le C4 en C4a et C4b, ainsi que le C2 en C2a et C2b
C4b s'associe avec C2a pour former C4b2a (une C3 convertase)
C4b2a clive jusqu'à 1'000 C3 en C3a et C3b
C3b s'associe avec C4b2a pour former C4b2a3b (une C5 convertase). Il peut aussi se lier spontanément à la surface du pathogène
Une fois formée, la C5 convertase va pouvoir cliver C5 en C5a et C5b, étape commune aux trois voies.
Voie alternative[]
voie alterne
La voie alternative est activée par de nombreux composants à la surface des bactéries, champignons et virus. C'est la première des voies mises en jeu lors d'une infection. La cascade de réactions est la suivante:
Une petite portion de C3 soluble s'hydrolyse spontanément en iC3 (une réaction qui se fait plus rapidement dans les milieux infectés à cause d'une modification de l'environnement par les constituants de certains pathogènes, en particulier des bactéries)
iC3 se lie au facteur B inactif, le rendant susceptible de se faire cliver par le facteur D
le facteur D clive le facteur B en Ba et Bb
Bb reste lié à iC3, formant le iC3Bb (une C3convertase)
iC3Bb clive des C3 en C3a et C3b
C3b se lie au pathogène
C3b sur le pathogène lie un nouveau facteur B, qui se fait cliver en Ba et Bb par un facteur D
le facteur Bb se lie avec C3b sur le pathogène, formant une C3bBb (qu'on appelle la C3convertasealternative) sur le pathogène
c3 convertase alterne
C3bBb clive juqu'à 1'000 C3 en C3a et C3b sur le pathogène, autour duquel se forme rapidement un manteau de complément
C3b peut s'associe avec C3bBb pour former C3b2Bb (laC5 convertase alterne). Il peut aussi se lier spontanément sur la surface du pathogène
C5 convertase alterne
Une fois formée, la C5 convertase va pouvoir cliver C5 en C5a et C5b, étape commune aux trois voies.
convergence des voies
Au final, le pathogène se retrouve rapidement enrobé de C3b à sa surface. La Properdine (ou facteur P), protéine du plasma, se lie à la C3 convertase alterne et la stabilise, augmentant sa demi-vie de 5min à 30min, en empêchant sa dégradation par des protéases.
MAC[]
MAC
Il y a une convergence des voies à partir de la formation de C5a et C5b par les diverses C5 convertases. Le C5b initie la formation de la MAC, ce qui va mettre en jeu C6, C7, C8 et C9. La MAC crée un trou dans la membrane du pathogène, avec un passage d'eau et de ions, capable de lyser certains pathogènes (particulièrement Neisseria meningitidis).
Le cellules humaines se protègent de la formation de MAC par la protéine S, la clusterine et le facteur J, qui empêchent la liaison de C5b, C6 et C7 avec la membrane cellulaire. Sur la surface cellulaire, le homologus restriction factor (HRF) et CD59 (protectine) empêchent le recrutement de C9.
Au final, la MAC a une utilité limitée comparée au large pouvoir d'opsonisation de C3b. (C4b est aussi capable d'osponisation).
Anaphylatoxines[]
anaphylatoxines
Les anaphylatoxines regroupent les C3a, C4a et C5a (C5a est le plus stable et le plus efficace). Elles peuvent induire la contraction du muscle lisse, la dégranulation des mastocytes et des basophiles, augmenter la perméabilité vasculaire ou encore induire le chimiotaxisme. De plus, C5a permet d'augmenter la capacité de phagocytose des neutrophiles et des macrophages et augmente l'expression de CR1 et CR3 sur leur surface. Ainsi, elles font une réponse inflammatoire locale, avec œdème et extravasation de protéines. Si la réponse est massive, elles peuvent avoir comme conséquence un choc anaphylactique, qui correspond à une réponse inflammatoire aigüe globale.
Récepteur au complément[]
Nos cellules possèdent des récepteurs aux complément. Un des plus important est le CR1, qui reconnait le C3b et le C4b.
récepteur
ligand
fonction
cellule
CR1
C3b, C4b
Phagocytose, nettoyage des complexes immuns
Érythrocytes, Macrophages, Dendritique folliculaire, PMN, Lymphocytes B
CR2 (CD21)
iC3b, C3b
C3d,
C3dg
Co-récepteur des lymphocytes B (signal 2 - rôle dans la production d'anticorps), récepteur à l'EBV
Lymphocyte B
Dentritique folliculaire
CR3
iC3b
Phagocytose
Macrophages, PMN, Dentritique folliculaires
CR4
iC3b
Phagocytose
Macrophages, PMN
En ce qui concerne les concentrationsanguines des protéines du compléments, les deux qu'on retrouve le plus sont le C3 surtout, et aussi le C4. Les autres se retrouvent à de très faible quantité dans le sang.
Régulation du complément[]
La régulation du complément se fait par plusieurs protéines, les Protéines RCA. Seul, le complément est une structure très instable. Une activation inappropriée du complément pourrait être dangereux pour l'organisme. Il existe plein de molécules régulatrices, certaines solubles, certaines sur des membranes.
facteurs P, H, I
Une molécule de régulation importante est le facteur H, qui se lie à C3b lié sur la surface du pathogène et facilite son clivage en iC3b (lié ≠ iC3b soluble qui fait partie de la voie alterne) par le facteur I (le facteur H est recruté par l'acide sialique, qui se trouve sur les cellules eucaryotes - mais les streptocoques et staphylocoque ont aussi des acides sialique et donc évitent l'activation du complément). Les morceaux d'iC3b liés ne peuvent pas assembler de convertase. Ainsi, l'action du facteur H et du facteur I s'oppose à l'action du facteur P (la Properdin).
Il existe plein de pathologies liées à des déficiences en complément. Par exemple la déficience en facteur H amènera à une formation continue de convertases, amenant à l'épuisement du stock en C3b. Les patient n'arrivent plus à correctement phagocyter les bactéries.
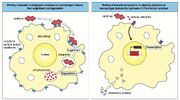
Phagocytose par les macrophages[]
phagocytose opsonisation
Les macrophages résiduels sont en général les premières cellules effectrices rencontrées par le pathogène. Les macrophages sont capables de phagocyter le pathogène. Le microbe se fait internaliser dans un phagosome (pH basique et activation enzymatique), qui fusionne avec un lysosome pour donner un phagolysosome (remplit de substances toxiques qui dégradent le microbe). Le processus de phagocytose est plus efficace si le pathogène possède des ligands qui se lient aux récepteurs de surface du macrophage. Un de ces récepteurs est le CR1, récepteur au C3b (qui opsonise le pathogène) déposés sur le pathogène par l'activation de la voie alternative du complément. Le CR1 protège également la membrane de la cellule où il est exprimé en facilitant le clivage de C3b et iC3b par le facteur I, afin de l'inactiver.
Il existe encore deux autres récepteurs aux molécules du complément. Le CR3 et le CR4, qui se lient aux fragments iC3b déposés à la surface du pathogène. Ensemble, les trois récepteurs sont les plus efficaces.
Structures propres aux pathogènes[]
Les microorganismes possèdent des structures stables dans l'évolution et différentes de celles de nos cellules, qu'on appelle desPAMP. Les cellules ont développé au fil de l'évolution des récepteurs, les PRR, pouvant reconnaitre les PAMP et initier la phagocytose (endocytose médiée par des récepteurs). Les Macrophages possèdent de nombreux récepteurs de ce genre, en plus de ceux pour le complément.
Une grande partie de ces structures étrangères sont des hydrades de carbones ou des lipides. Les récepteurs et protéines plasmatiques capables de reconnaitre les hydrates de carbones sont appelés les lectines (par exemple le récepteur MBL, ou le récepteur au glucan).
Les macrophages possèdent aussi un récepteur nommé scavenger, qui peut reconnaitre toute une panoplies de structures qui possèdent le point commun d'être négativement chargées (comme des polysaccharides sulfatés, des acides nucléiques, des LTA, etc.).
Les récepteurs du complément CR3 et CR4, en plus de lier le iC3b, peuvent aussi reconnaitre le LPS des bactéries Gram- , ainsi que d'autres structures spécifiques à certains pathogènes (dont ils stimulent la phagocytose).
PRR[]
En plus de pouvoir phagocyter, les macrophages possèdent aussi des récepteurs leur permettant de sécréter des cytokines inflammatoires (indispensables pour la réponse innée et pour la réponse adaptative si elle est nécessaires) lors de présence de pathogènes, qui vont recruter d'autres cellules immunitaires dans les tissus.
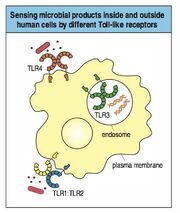
Une des familles de récepteurs les plus importants dans ce domaine sont les TLRs(les TLRS reconnaissent des PAMP, qui sont absents des cellules de mammifères, mais présents sur beaucoup de pathogènes), qui induisent la production de cytokines.
TLRs
Chaque TLR est spécifique à un type de microbes différents, ce qui permet de cibler la réponse immunitaire. On les retrouve sur les cellules dendritiques, les macrophages et les cellules B notamment.
Le macrophage exprime le TLR4 qui est spécifique pour leLPS bactérien. Lorsqu'il reconnait le LPS, le TLR4 va activer dans le noyau du macrophage les gènes permettant de synthétiser les cytokines inflammatoires (via le facteur de transcription NFkB). En plus du LPS, le TLR4 peut reconnaitre d'autres structures bactériennes qui lui ressemblent, ainsi que des structures fongiques (mannans).
L'homme possède 10 TLRs différents, qui peuvent être des homodimères ou des hétérodimères. Chacun reconnait des familles de structures différentes (double brin d'ARN, LTA, Flagelline, Zymosan, etc.). Ensembles ils sont capables de reconnaître un très grand nombre de pathogènes.
ADN CpG (=coiffé) non-méthylé bactérien et viral: TLR9
ARN viral double brin: TLR3
ARN viral simple brin: TLR7 et TLR8
Les TRLs se retrouvent soit à la surface de la cellule, soit à l'intérieur des endosomes (pour reconnaitre les acides nucléiques - TLR3, TLR7, TLR8, TLR9).
Différents récepteurs TLR
Souvent, les TRLs ne sont pas les seuls à reconnaitre le pathogène, mais fonctionnent comme des co-récepteurs. Par exemple, LPS est co-lié par le CD14 et le TLR4 (qui est plutôt le senseur que le lieur).
Les protéines NODsont un autre groupe de récepteurs intracellulaires reconnaissant les composés peptidoglycans bactériens (une mutation de NOD peut être impliquée dans la maladie de Crohn).
Le RIG-1est un récepteur cytoplasmique qui reconnait les ARN double brin des virus. Il déclenche la production d'IFN-I (qui inhibe la réplication virale et cible la réponse immunitaire en recrutant Th1)
Les MBLreconnaissent les mannoses typiques (PAMP) des parois bactériennes. On les trouve sur les macrophages et les neutrophiles (récepteur au mannose) ou sécrété (MBL) permettant la phagocytose et l'activation de la voie des lectines du complément.
Les scavengerrécepteur reconnaissent les ligands chargés négativement, spécifiques des bactéries Gram+. Il est surtout présent sur les macrophage et stimule la phagocytose.
Le récepteur aux peptides N-formylés est aussi un PRR.
Cytokines[]
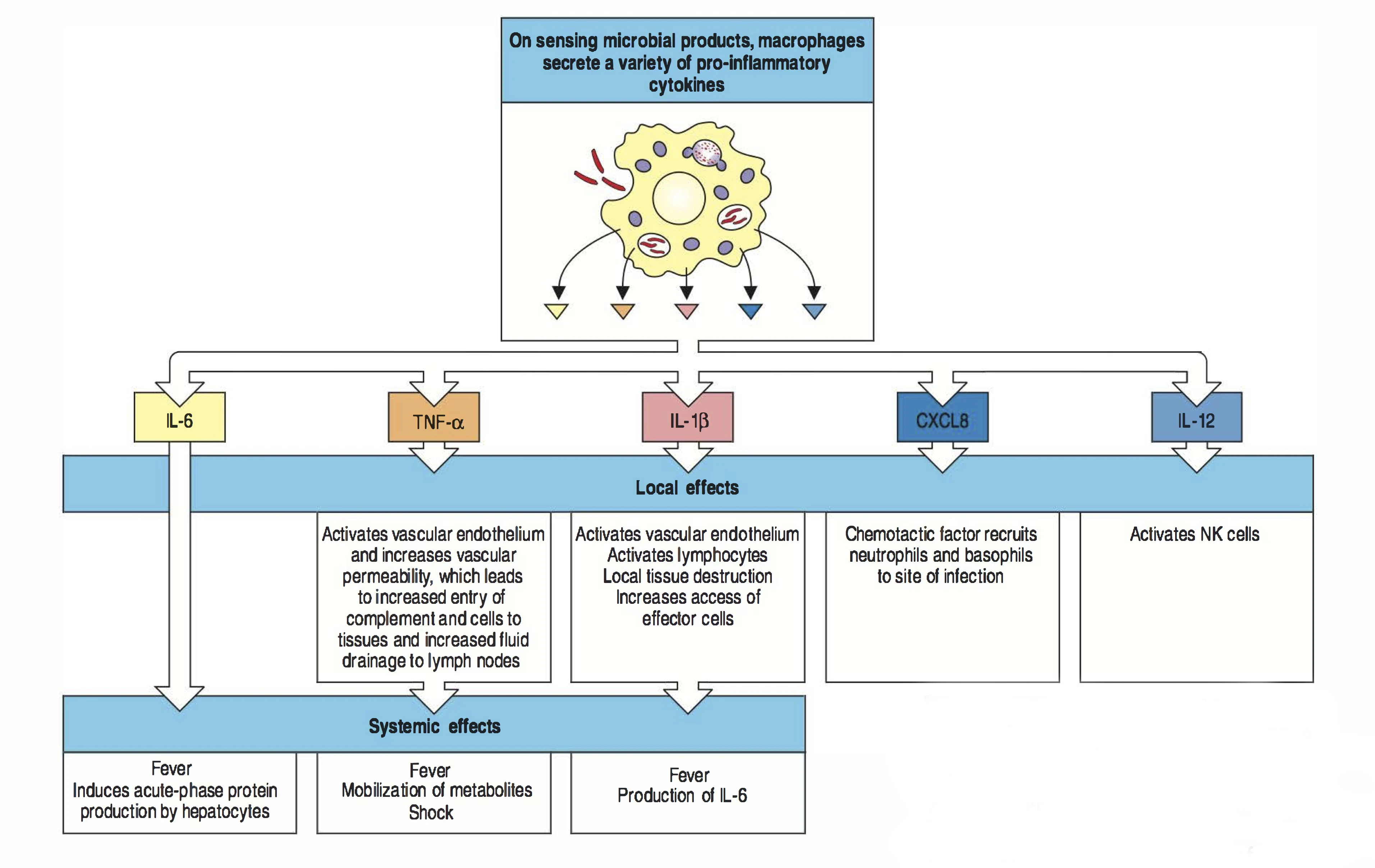
cytokines macrophage
Lorsque le macrophage répond à la reconnaissance d'un pathogène, il produit une variété de cytokines. Les cytokines sont des petites protéines qui vont aller se lier à la surface d'autres cellules et induire une réponse locale ou systémique. Les cytokines importantes produites par le macrophage sont:
IL-1
IL-6
CXCL8
IL-12
TNF-alpha
Ces cytokines possèdent de puissants effets qui peuvent être locaux ou systémiques.
CXCL8 (ou IL-8), est une chémokine. Les chemokines ont comme effet d'attirer d'autres cellules. CXCL8 attire les neutrophiles au site d'inflammation. Les neutrophiles possèdent deux récepteurs aux CXCL8: le CXCR1 et le CXCR2. Lorsqu'ils sont activés, les neutrophiles sont structurellement altérés de manière à être capable de migrer dans les tissus. Le chemin les guidant jusqu'au site d'infection se fait grâce à un gradient de chemokines présente en solution et aussi attachées sur la matrice et sur les cellules endothéliales. Certaines chémokines ont une activité anti-microbienne.
IL-12 a comme fonction d'activer les cellules NK. Les NK sont spécialistes pour les infections de types virales.
IL-1 et TNF-alpha (réponse inflammatoire locale) ont des effets sur l'endothélium permettant de faciliter l'entrée des cellules effectrices sur le site d'infection. De plus, IL-1, IL-6 et TNF-alphastimulent la synthèse de protéines de la phase aiguë par le foie et augmentent la température corporelle.
TNF-alpha induit également la production de PAF par les cellules endothéliales, qui induit le blocage des vaisseaux par formation de caillots, ce qui limite l'entrée du pathogène dans la circulation systémique. Si l'action du TNF-alpha (vasodilatation, augmentation de la perméabilité, système de PAF,...) se produit de façon systémique lors d'infection du sang, cela mène à un choc septique.
Nomenclature[]
Interféron: protéine antivirale
Interleukine: médiateur entre les lymphocytes
Lymphokine: médiateur produit par les leucocytes
Monokine: médiateur produit par les monocytes
Cytokines type 1 et 2
Chémokine: molécule chémotactique
Adipokine: molécule impliquée dans le métabolisme
Facteur de croissance et de différenciation: pour les cellules hématopoïétiques, neuronales etc.
Cytokines de type 1: IFN-gamma et IL-12, qui sont produites par ou induisent les cellules Th1
Cytokines de type 2: IL4, IL5, IL13, qui sont produites par ou induisent les cellules Th2
Rôles[]
Prolifération et différenciation cellulaire
Fibrose
Hématopoïèse
Immunité
Inflammation
Réparation tissulaire
Réseau neuroimmun
Cancer
Athérosclérose
Pathogenèse HIV-1
Neutrophiles[]
neutrophile vs bacterie
Les neutrophiles possèdent plein de récepteurs contre les pathogènes, ainsi que des récepteurs pour le complément pour faciliter la phagocytose.
Comparés aux macrophages, les neutrophiles ont une plus grande variété d'action, une plus grande variété de substances microbicides, mais une durée de vie beaucoup plus courte. Ils sont programmés pour mourir jeune. L'arrivée des neutrophiles est la première étape de la réponse inflammatoire.
Les phagosomes des neutrophiles vont fusionner avec les deux types de granules qu'ils possèdent. Les granulesprimaires (ou azurophiles) et les granulessecondaires (ou spécifiques). Les granules primaires sont remplis de protéines pouvant digérer les microbes (lysozymes, défensines, myélopéroxydases, élastases, protéases, ...). Les granules secondaires contiennent entre autres la lactoferrin, qui fait compétition avec le pathogène pour le fer et le cuivre, et la NADPH oxydase, qui produit des ROS qui seront convertis en H2O2, via la SOD. Cela aura comme effet d'activer les enzymes des granules du neutrophile qui vont pouvoir efficacement attaquer le pathogène.
Après avoir fusionnés avec les granules, les phagosomes vont fusionner avec les lysosomes contenant des hydrolases acides pour former les phagolysosomes.
L'augmentation des ROS (qui s'accompagne d'une augmentation de la consommation d'oxygène appelée respiratory burst) est accompagnée d'une production de catalase pour éviter d'endommager les cellules de l'hôte.
Le neutrophile, une fois dégranulé, ne peut recréer les granules et s'apoptose, avant d'être phagocyté par les macrophages.
Granulomatose septique[]
granulomatose septique
La granulomatose septiquechronique (chronic granulomatous disease) est une maladie se caractérisant par un défaut génétique du gène codant pour la NADPH oxydase.
Sans NADPH oxydase, les enzymes du PMN ne peuvent pas s'activer, et celui-ci peut phagocyter mais pas détruire le pathogène (il n'y a pas de respiratory burst avec production de O2- et le pH ne peut pas être augmenté pour activer les enzymes nécessaires à la destruction du pathogène). Les bactéries et champignons peuvent donc proliférer.
L'infection sera contenue par les autres cellules immunitaires dans des granulomes contenant des macrophages remplis de neutrophiles.
Parmi les bactéries qui sont présentes dans cette maladie, on retrouve notamment E.coli. Parmi les champignons, on retrouve notamment Aspergillus fumigatus.
La granulomatose septique chronique est une maladie génétique liée à l'X dans 60% des cas (et autosomale récessive dans le reste des cas).
Le traitement inclut:
Antibiotiques contre les infections bactériennes
Amphotéricine B contre l'Aspergillus fumigatis
IFN-gamma à long-terme, pour stimuler les macrophages et les NK (elle augmente aussi la présentation des antigènes)
Le meilleur traitement est la greffe de cellules hématopoïétiques
Les déficits en G6P-DH ou en myéloperoxydase peuvent faire des symptômes similaires. La leukocyte adhesion deficiency est une mutation de LFA-1 qui empêche la fixation de I-CAM à l'endothélium et empêche la diapédèse. Le syndrome de Chédiak-Higashi empêche la fusion des vésicules avec le lysosome.
Phase aigüe[]
IL-1 IL-6 TNF-alpha
Les cytokines IL-1 , IL-6 et TNF-alpha ont la capacité d'induire de la fièvre. La fièvre est une augmentation de la température du corps. Ces cytokines agissent au niveau des centres de contrôle de la température dans l'hypothalamus, et dans les muscles et cellules adipeuses (en altérant la mobilisation d'énergie, afin de générer de la chaleur). Les molécules qui induisent la fièvre sont nommées "pyrogènes endogènes". Certains pathogènes possèdent des molécules pyrogènes (molécules pyrogènes exogènes).
La fièvre est utile pour la lutte contre les pathogènes, car souvent ceux-ci sont plus prolifératifs à de basses température. De plus, la fièvre diminue les effets néfastes de TNF-alpha sur les cellules, qui sont plus résistantes à haute température.
phase aigue
Un autre effet des IL-1, IL-6 et TNF-alpha est la production de protéines de la phase aigüe par les hépatocytes. Parmis celles-ci, on retrouve la MBL , la CRP et le fibrinogène.
La MBL, de la famille des collectines, est augmentée lors de la phase aiguë en forme soluble et va se lier aux mannoses des pathogènes (bactéries, champignons, protozoaires et virus) et activer la voie de la lectine du complément. Elle permet aussi d'opsoniser le pathogène
La CRP, de la famille des pentraxine, va se lier à la phosphocholine (un composant des lipopolysaccharides de la paroi des bactéries et champignons) et activer la voie classique du complément. Elle permet aussi d'opsoniser le pathogène.
Hors des infections, les protéines de la phase aiguë sont présentes dans le plasma mais à de très petites quantités. Lors des infections, leur quantité augmente de 1'000x pendant le pic de la réponse aiguë (à 2j après le début).
Notes[]
Une cellule qui finit par "blaste" est un précurseur.


























































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


{kind=link}


{kind=link}


{kind=link}


{kind=link}